
Latitudinal Diversity Gradient
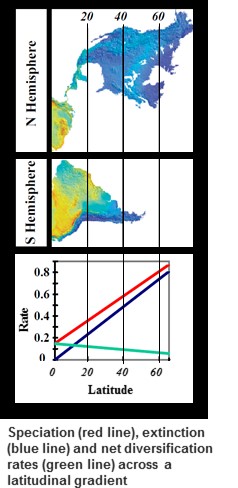
A growing number of studies have demonstrated a higher net rate of diversification (species origination rate minus extinction rate) in the tropics compared to temperate regions. A prevalent view is that accelerated tropical net diversification rates result from faster species originate rates there. Alternatively, low rates of extinction could also drive higher net diversification rates, a concept put forward by the 19th century explorer and biogeographer Wallace. To test these alternative hypotheses I compared the distribution of sister species ages in temperate versus tropical birds and mammals of the New World. Age distributions were pushed up towards the present at high latitudes (i.e. were young) but extended much further into the past in the tropics (i.e. were older). Part of this pattern may be generated by taxonomic uncertainty with tropical species appearing older simply because we have yet to define them properly. However, the relationship of decreasing age with latitude remains significant within North America where species boundaries are well studied. Using a model that incorporates taxonomic uncertainty (and thus corrects for a possible taxonomic artifact in the tropics) I compared the empirical distributions of sister species ages to ones simulated under a variety of parameters of speciation and extinction. Maximum likelihood methods were used to determine which parameters of speciation and extinction produced sister species age distributions that best matched the empirical dataset of sister species ages (Fig. 1). Key results are as follows (
Weir & Schluter 2007
):
1) rates of species origination increased significantly with latitude (Fig.1) in direct opposition to theory that accelerated tropical rates drive the excess of species.
2) extinction rates also increased significantly with latitude
3) species turnover was much higher at high latitudes than in the tropics supporting Wallace’s old notion that stability of the tropics resulted in low extinction and allowed for the gradual accumulation of high levels of species diversity
These results support the key role of the tropics as a "museum" preserving diversity from extinction and allowing a gradual accumulation of high diversity. In contrast, high latitudes act as a "cradle" of diversity, but loose this diversity rapidly through extinction.
Current projects estimate rates of evolution in species recognition traits (color, song etc.) to determine if they also evolve faster towards the poles.
Great American Biotic Interchange
North and South America have been island continents for most of the past 90 million years until completion of the Central American Landbridge connected the continents about 3.5 million years ago. During its isolated island phase South America evolved highly distinctive mammalian, avian and other faunas in what the palaeontologist Simpson referred to as "splendid isolation". Following this long episode of isolation, the fossil record reveals a massive wave of interchange between North and South America as mammals freely roamed between the continents. However, the fossil record is very poor for birds. Given their ability to fly across water barriers it was unknown if landbridge completion also facilitated a massive wave of interchange in birds. Where fossils fail, my collaborators and I used DNA sequences and a molecular clock to date interchange events between North and South America in four large Neotropical families and a number of smaller ones. An example phylogeny of the forest specializing woodcreepers with maximum likelihood ancestor state reconstruction of occurrence in North or South America is shown in Fig 2. This reconstruction shows a close correlation between landbridge formation and the first interchange event of woodcreepers from South America into North America. Antbirds showed similar results demonstrating the need for a completed landbridge in rainforest restricted avian families. In contrast the more habitat generalist tanagers and blackbirds have each colonized isolated oceanic islands and their reconstructions showed less reliance on a completed landbridge. These results suggest that continental interchange in rainforest restricted avian groups, like most mammals, was tied closely to a completed landbridge.

Paleoclimate Change and Diversification

Climatic fluctuations may play a key role in diversification in several ways. First, climatic fluctuations may speed up the evolutionary process by increasing the opportunity for allopatric vicariance and thus speciation. Second, shifting environmental factors may result in an ever-changing selection regime, accelerating the rate at which populations adapt and change. Third, climatic fluctuations may increase extinction risks. I am investigating the role of climatic fluctuations on diversification at both temperate and tropical latitudes.
In North America, I compared ages of species belonging to superspecies complexes in the high latitude boreal zone with temperate species south of the boreal zone. Boreal species belonging to superspecies complexes were younger and each date to the Pleistocene glacial periods. These high latitude species complexes share a common pattern of endemism with species endemic to the Taiga east of the Rockies, to coniferous forest in the Rocky Mountain region and to mesic coniferous forests along the Pacific Coast. A common pattern of phylogenetic branching supports a sister relationship between the Pacific and Rocky Mountain species with a more distant relationship to Taiga species. Dates of splitting correlate with key features of the Pleistocene glacial cycles (Fig. 3). While almost all boreal superspecies complexes date the Pleistocene, many sub-boreal and tropical lowland superspecies complexes predate the Pleistocene. These differences in superspecies ages suggest faster rates of evolutionary and ecological divergence at high latitudes where climatic fluctuations have been most intense.

In the Neotropics, I examined the ages of evolutionary splitting events between species level taxa (i.e. nodes) in phylogenetic trees (Fig 4). I compared ages of species level nodes in highland genera whose geographic ranges were heavily impacted by glacial cycles during Pliocene and Pleistocene with lowland genera where climatic perturbations accompanying ice ages had minimal impact. Not only are species level nodes younger on average at high altitudes of the Neotropics, but lineage through time analysis indicates that most highland genera have maintained a fairly constant rate of diversification up the present. In contrast species in most lowland genera are old and lineage through time analysis indicate a significant fauna wide decline in speciation rates towards the present consistent with density dependant cladogenesis. These results suggest that opportunities for ongoing speciation in lowland faunas may have slowed through time while diversification rates remain steady in highland regions where climatic fluctuations and ongoing orogeny may create new opportunities for further diversification. One caveat to these results is that many lowland species may contain additional phylogenetic splits (i.e. phylogroups) which are not recognized as species under the biological species concept but would be under the phylogenetic species concept. Inclusion of phylogroup splits in diversification analysis will add additional young nodes to lowland faunas. However, even if we consider every lowland subspecies to represent a distinct lineage in these analysis many lowland genera would still exhibit a slow down in diversification rates through time (Fig 5).

Molecular Clocks

If DNA mutations accumulate at a more or less constant rate through time, then it should be possible to use the number of mutations that differentiate two populations to obtain a date at which they split from a common ancestor. To achieve such dating, calibration of the rate of mutation accumulation is necessary. A few molecular clocks have been generated for birds using fossils and biogeographic events. These have generally provided a rate close 2% per million years (1% per lineage) in protein-coding mitochondrial DNA. The 2% calibration has been used widely to date the origin of taxonomic groups, date key dispersal events and calibrate rates of speciation and extinction. However, the 2% clock is based on only a handful of calibration events and levels of rate variation across different orders of birds is unknown.
To address these uncertainties, I focussed on obtaining a dataset of 90 potential molecular clock calibrations for birds (Weir & Schluter 2008). These were obtained either from dated fossils or from dated biogeographic events (e.g. dates of islands, mountains or land bridges). A cross validation method (see CCVAL 1.0 in software) was utilized to discriminate between genuine rate variation across lineages and faulty or highly imprecise calibration points that artificially inflate measures of rate variation. The key findings are as follows:
1) corrected genetic distances accumulate with time (up to about 12 million years) in a linear fashion (Fig 6)
2) average molecular rates were 2.1%
3) average molecular rates did not vary greatly across taxonomic orders
4) rate variation across lineages was present but was not explained by differences in generation time, body mass or latitude as previously suggested. Rather fossils tended to overestimate molecular rates and biogeographic events underestimated molecular rates significantly explaining some of the observed rate variation
Phylogeography and Phylogenetics
Projects are underway investigating the molecular phylogenetics, taxonomy and biogeography of several New World (Chlorospingus bush-tanagers and Dendrocincla woodcreepers) and Old World genera using a variety of molecular, vocal, morphological and plumage based analyses.