
Welcome to the Weir Lab at the University of Toronto. Our lab uses genomic and comparative methods to investigate the pace of evolution in the tropics versus high latitudes. The tropics have exceptional levels of biodiversity, and understanding the drivers of that diversity is an underlying theme in our lab’s research. As lab members we are also committed to sustainable lifestyle choices that are necessary for the biodiversity we study to thrive on our planet (see Biodiversity and Sustainable Living). Using birds as our model group, we employ a variety of genomic, field and computational approaches to estimate evolutionary rates across latitudinal gradients.
The Weir Lab is proud to collaborate with excellent colleagues in New Zealand, Brazil, and Israel (Hebrew University).
LAB AWARDS
Hybrid Speciation in Amazonian Birds

The Genomics of Speciation
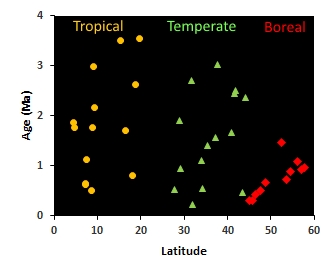
A recent focus of the lab is on the genomics of speciation, and how the genetic architecture of reproductive isolation varies between birds living in the rainforests of the Amazon versus the boreal forests of Canada. Our past work has shown that closely related bird species in the tropics continue to form hybrid zones for up to four million years following divergence (see figure on left), while the oldest hybrid zones at boreal latitudes are less than 1.5 million years old (read.here). The older age of hybridizing species in the tropics suggests that tropical species are accumulating reproductive isolation more slowly there, and thus taking a longer time to complete

Phylogenetic estimates of speciation and extinction rates
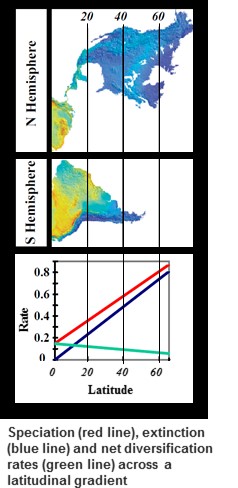
Are speciation rates fastest in the tropics, and can they explain the high species richness there? Using phylogenetic data we showed that rates of speciation have been much faster at high latitudes, and slower in the tropics over the past few million years. In contrast, extinction rates were very slow in the tropics, and might explain how the tropics have been able to accumulate such high levels of species richness (read here). Recent research by former Ph.D. student Dr. Paola Pulido-Santacruz extended this work across the entire bird phylogeny. Her work suggests that extinction rates are consistently lower in geographic regions with low species richness across many replicate orders of birds, while speciation rates showed no consistent pattern with latitude across replicate clades (read here). Together, these studies suggest that low tropical extinction could be the key historical driver behind the latituidnal diversity gradient.
Latitudinal gradients in trait evolution
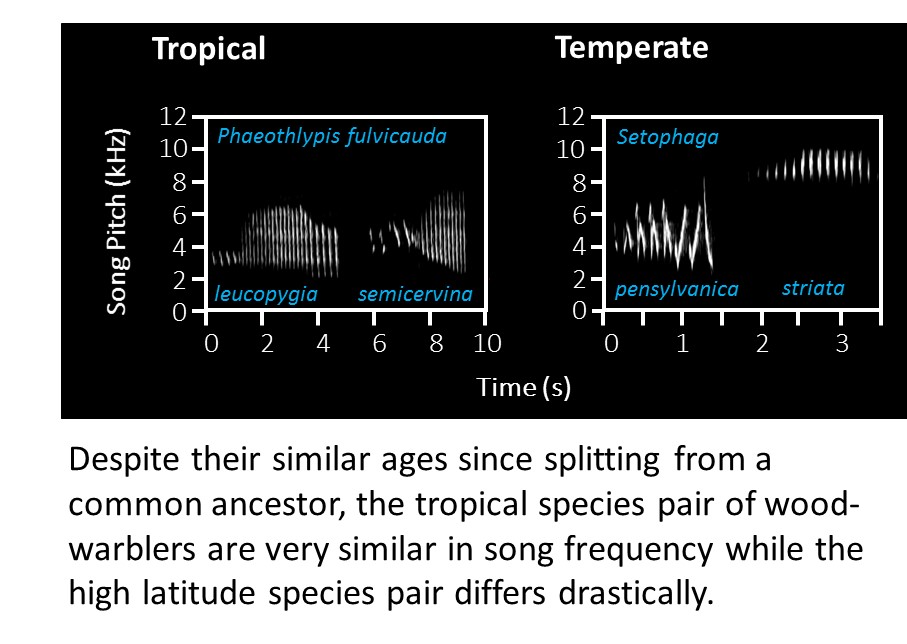
We investigate whether rates of evolution vary across latitudinal and ecological gradients in traits (song, colour, morphometrics and climatic niche) important for speciation and ecological differentiation. A key focus has been on rates of evolution in the courtship songs used by birds to attract mates. Our results demonstrate faster evolution of bird song (Weir & Wheatcroft 2011, Proc. Roy B; Weir et al. 2012 Evolution. Former MSc student Adam Lawson demonstrated a much faster rate of climatic niche evolution at high latitudes, suggesting that divergent selection pressures are stronger towards the poles (Lawson & Weir 2014, Ecology Letters) while PhD student Vanessa Luzuriaga-Aveiga has demonstrated that increased elevational differentiation results in increased rates of song and morphometric trait evolution across the Amazon and eastern Andean slopes (Luzuriaga-Aveiga & Weir 2019, Ecology Letters). PhD student Sean Anderson has developed new models of trait evolution (and a new R package) that quantify the degree of divergent adaptation (Anderson & Weir 2020, Am. Nat.) and more recently that distinguish between character displacement,
Comparative Phylogeography and Biogeography

An ongoing focus of the lab is to utilize comparative approaches across many species to address key biogeographic questions such as the role of ice sheets in driving allopatric speciation in boreal birds (read.here) and New Zealand kiwi (read here), the role of climate change in driving diversification of lowland and highland birds of the Neotropics (read here), and the role of landbridge completion in driving the Great American Biotic Interchange between North and South American birds (read here). We are using genome-wide and whole genome datasets to study phylogeographic questions in a variety of other North American and Amazonian species: manakins (Ph.D. student Else Mikkelsen and former student Dr. Alfredo Barrera-Guzman), antbirds (former students Dr. Aurea Cronemberger and Dr. Maya Faccio), tanagers (Ph.D. student Vanessa Luzuriaga), flyctachers (former post doc Dr. Jordan Bemmels), warblers, thrushes (with collaborators Alyssa FitzGerald and Jeremy Kirchmam), titmice (with collaborator Claire Curry), and kiwi (with former post doc Dr. Jordan Bemmels).
Weir Lab Opportunities (details here)
